Background
Lateralization disorders are divided into complete (ie, situs inversus totalis) and incomplete (ie, heterotaxy); the word heterotaxy is derived from the Greek heteros, meaning “other” and taxis, meaning “arrangement.” The disorders have been recognized since at least 1933 (complete) [1] and 1826 (incomplete). [2] Only relatively recently have genetic alterations responsible for their occurrence in humans been identified. The discovery of kindreds in which both heterotaxy and situs inversus totalis occur strongly suggests that these are not truly separate diseases. [3] At least 12% of primary ciliary dyskinesia (PCD) patients have heterotaxy. [4]
Because PCD is present in about a quarter of situs inversus totalis patients and due to the fact that situs inversus totalis is present in nearly half of individuals with PCD, this article covers both heterotaxy syndrome and PCD. Moreover, because asplenia and polysplenia can occur in the same family, [5, 6] a patient’s splenic phenotype should be viewed as merely one phenotypic aspect of an underlying laterality disorder, even though many prior reviews of heterotaxy syndrome have divided patients into two groups based on only splenic phenotype.
Pathophysiology
Embryology and developmental biology
Ivemark’s review of 65 cases of human patients with asplenia, [7] in which most but not all had heart disease, firmly established the spectrum of congenital heart lesions that occurred in patients with lateralization disorders. Four years later in 1959, the recovery of a spontaneous, autosomal recessive, viable mutation in mice was reported and was named iv, for inverted viscera. [8]
Although the stomach position in the iv mutant colony remained perfectly randomized over 15 years of breeding, including only one outcross, the prevalence of discordance between thoracoabdominal venous anatomy and the situs of the rest of the body decreased from 42% to 26%. [9] That is, the ratio of the heterotaxy phenotype to the situs inversus totalis phenotype decreased with progressive inbreeding. This suggests that the wild type allele of the iv locus controls overall thoracoabdominal sidedness and not individual organ sidedness. Indeed, when Icardo and Sanchez de Vega examined the hearts of iv homozygotes, only 40% were abnormal, and only 36% had abnormal splenic morphology. [10, 11]
The absence of outer dynein arms in the spermatozoa and airway cilia of humans with the Kartagener triad (ie, situs inversus totalis, sinusitis, and bronchiectasis) was also noted by Björn Afzelius (see the image below).
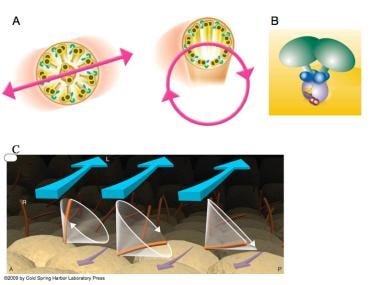 The structure and function of cilia is shown here. (A) Most motile cilia are organized with 9 microtubule doublets surrounding a core pair of doublets (9+2 configuration). Outer dynein arms (green) and inner dynein arms (blue) are shown. Cilia on the cells of the ventral node in the normal mouse embryo have no core doublet (a 9+0 configuration) and were initially thought to be nonmotile; however, upon closer scrutiny, node cilia were seen to have a rotatory motion (600 rpm). [Figure A is from Hirokawa N, Tanaka Y, Okada Y. Left-right determination: involvement of molecular motor KIF3, cilia, and nodal flow. Cold Spring Harb Perspect Biol. Jul 2009;1(1):a000802 and is reprinted with permission of Cold Spring Harbor Press.] (B) lrd (left-right dynein), the protein (green) mutated by the iv mutation, is also known as DNAH11, DNAHC11, and DLP11. [Figure B is from the United States Department of Energy Genomes to Life Program.] (C) The rotatory cone of each cilium is tilted posteriorly. Hence, the cilia make a leftward swing at the fluid surface and a rightward swing at the cellular surface. Because more viscous drag is present at the cellular surface, the rightward sweep is less effective at generating fluid movement than is the leftward sweep. [Figure C is from Hirokawa N, Tanaka Y, Okada Y, Takeda S. Nodal flow and the generation of left-right asymmetry. Cell 2006; 125:33-45 and is reproduced with permission from Cell Press.] A = anterior; L = left; P = posterior; r = Right.
The structure and function of cilia is shown here. (A) Most motile cilia are organized with 9 microtubule doublets surrounding a core pair of doublets (9+2 configuration). Outer dynein arms (green) and inner dynein arms (blue) are shown. Cilia on the cells of the ventral node in the normal mouse embryo have no core doublet (a 9+0 configuration) and were initially thought to be nonmotile; however, upon closer scrutiny, node cilia were seen to have a rotatory motion (600 rpm). [Figure A is from Hirokawa N, Tanaka Y, Okada Y. Left-right determination: involvement of molecular motor KIF3, cilia, and nodal flow. Cold Spring Harb Perspect Biol. Jul 2009;1(1):a000802 and is reprinted with permission of Cold Spring Harbor Press.] (B) lrd (left-right dynein), the protein (green) mutated by the iv mutation, is also known as DNAH11, DNAHC11, and DLP11. [Figure B is from the United States Department of Energy Genomes to Life Program.] (C) The rotatory cone of each cilium is tilted posteriorly. Hence, the cilia make a leftward swing at the fluid surface and a rightward swing at the cellular surface. Because more viscous drag is present at the cellular surface, the rightward sweep is less effective at generating fluid movement than is the leftward sweep. [Figure C is from Hirokawa N, Tanaka Y, Okada Y, Takeda S. Nodal flow and the generation of left-right asymmetry. Cell 2006; 125:33-45 and is reproduced with permission from Cell Press.] A = anterior; L = left; P = posterior; r = Right.
When the iv mouse mutation was cloned, it was found to encode a molecular motor protein, an axonemal dynein, and was named lrd, for left-right dynein (human homolog is DNAH11/DNAHC11, Dynein heavy chain 11, axonemal). However, its expression at embryonic day 7.5 was confined to the few hundred ciliated cells of the ventral surface of the node, a fluid-covered, pit-shaped structure at the anterior end of the primitive streak. Because these cilia, 5 microns in length and 0.3 microns in diameter, are missing the central doublet (i.e., have a 9+0 configuration of microtubule doublets, rather than the 9+2 configuration typically seen in motile cilia), they were not believed to be motile. Although the node was known to have important roles in organizing the body plan of the mouse embryo, the function of lrd remained mysterious.
The next year, the phenotype of mice missing the kinesin Kif3b was reported. Kif3b is a molecular motor which, like dynein, is responsible for transport along microtubules within cilia; kinesins transport their cargo toward the "plus end" of the microtubule, whereas dyneins are "minus end directed motors." Fifty percent of the 9.5-day embryos had L-looped hearts. Closer scrutiny of the cilia on normal ventral node cells showed that they do in fact move, despite their 9+0 arrangement of microtubules. In fact, uniquely among cilia, they rotate at 600 rpm. Ventral node cells of Kif3b nulls had either sporadic, very short cilia or absent cilia. [12] Whereas iv heterozygotes had cilia that rotated at 600 rpm, iv homozygotes had immotile cilia.
Because of a posterior tilt in the orientation of the cilia, as well as a difference in the viscous drag at the fluid surface compared with the base of the pit, the fluid within the node pit moves unidirectionally to the left, as verified by the movement of submicron-sized fluorescent beads applied to the fluid as passive tracers. [13] This fluid flow sets up a left-right asymmetric distribution of signaling molecules (eg, the evolutionarily conserved nodal/Pitx2 pathway—specifying the right side in protostomes and nonchordate deuterostomes but the left side in chordates [14] ) within the embryo during gastrulation, when the three germ layers (ie, ectoderm, mesoderm, and endoderm) are specified. Although some investigators initially proposed that perinodally situated sensory cilia might transduce the force of fluid flow via intracellular calcium signaling, mechanical forces do not in fact evoke measurable increases in intracellular calcium. [15]
In the zebrafish (Danio rerio), rotatory cilia-bearing structures homologous to the mouse node have been identified; however, they do not appear to be present in chicks or pigs, [16] which both appear to have short, nonmotile cilia [17] and may utilize an alternative left-right specification strategy at the embryonic node. [18] Moreover, the African clawed frog (Xenopus laevis) specifies the embryonic left-right axis long before cilia can be identified. Numerous steps in the process of embryonic development that precede the development of node cilia remain unknown. In fact, three temporal phases in which molecular and cellular decisions determine the left-right axis of the body plan are likely (ie, pregastrulation, gastrulation, and organogenesis), as is shown in the image below.
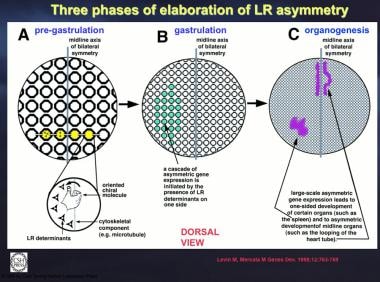 Three phases of elaboration of left-right (LR) asymmetry are shown. The first step consists of differentiating the left and right sides on the cellular level. This probably takes place by means of a chiral molecule. (A) A subset of the cells (yellow) of the fairly early embryo undergo this process. (B) Localized cellular asymmetry is propagated between cells to cause LR determinants to accumulate on one side of the embryonic midline, possibly by a process involving transport through gap junctions. These determinants would then induce cascades of factors in multicellular fields of the embryo. (C) Finally, the asymmetric presence of these factors induces or suppresses asymmetrically located organs such as the spleen and regulates asymmetric morphogenesis of other organs such as the heart tube. Courtesy of Levin M, Mercola M. The compulsion of chirality: toward an understanding of left-right asymmetry. Genes Dev. Mar 15 1998;12(6):763-9.
Three phases of elaboration of left-right (LR) asymmetry are shown. The first step consists of differentiating the left and right sides on the cellular level. This probably takes place by means of a chiral molecule. (A) A subset of the cells (yellow) of the fairly early embryo undergo this process. (B) Localized cellular asymmetry is propagated between cells to cause LR determinants to accumulate on one side of the embryonic midline, possibly by a process involving transport through gap junctions. These determinants would then induce cascades of factors in multicellular fields of the embryo. (C) Finally, the asymmetric presence of these factors induces or suppresses asymmetrically located organs such as the spleen and regulates asymmetric morphogenesis of other organs such as the heart tube. Courtesy of Levin M, Mercola M. The compulsion of chirality: toward an understanding of left-right asymmetry. Genes Dev. Mar 15 1998;12(6):763-9.
For technical reasons, the pregastrulation time period is particularly difficult to study in mammals. Important left-right axis specification decisions may occur in this developmental time interval in mouse and human; thus, the frog, chick, and pig may not be “outliers.”
In addition, the underlying cellular biology of why improper left-right specification of the lateral plate mesoderm has such a profound effect on the patterning of the heart, particularly the venous inflow and arterial outflow, has yet to be understood.
Anatomy
Predominantly endodermal structures
The bronchial branching pattern (and lung lobation) can be normal, inversus, right isomeric, or left isomeric. Liver lobation can be normal, inversus, or symmetric. In the gallbladder and biliary tree, hypoplasia, absence, and duplication can be noted. The spleen can be normal, absent, hypoplastic, or multiple. The intestine can develop malrotation.
Predominantly mesodermal structures
The hepatic segment of inferior vena cava (IVC) can be present or absent (the so-called “interrupted IVC"). The hepatic veins can be normal (ie, join the IVC just proximal to the IVC-atrial junction) or can connect independently to the atria. The coronary sinus can be normal, absent, or completely unroofed. The superior vena cava (SVC) can be normal (unilateral) or bilateral. Pulmonary veins can be partially anomalous or totally anomalous. Appendage morphology can be normal, inversus, right isomeric, or left isomeric. The common atrioventricular canal (CAVC) is usually significantly malaligned toward the morphologic right ventricle (RV), but it can be malaligned toward the morphologic left ventricle (LV). The ventricles can be D-loop or L-loop (superoinferior ventricles is rare). In the outflow tract, subpulmonary stenosis or atresia is usually noted, but subaortic stenosis can be observed. Double-outlet RV is most common, but tetralogy of Fallot can occur. Transposition of the great arteries can occur.
Either a left aortic arch with a left upper descending aorta or a right aortic arch with a right upper descending aorta can occur. Double aortic arch is exceedingly rare. In cases of left aortic arch with left upper descending aorta, the abdominal aorta is left of the spine. In cases of right aortic arch with right upper descending aorta, the abdominal aorta is right of the spine (unlike the situation without heterotaxy, in which the abdominal aorta is left of the spine). Many cases have both the abdominal aorta and the IVC (or azygos, if the hepatic IVC is absent) on the same side of the spine (unlike without heterotaxy, in which the IVC is right of the spine, whereas the abdominal aorta is left of the spine).
Although the genetic underpinnings of left-right patterning of the embryonic brain and spinal cord have been studied extensively in some vertebrate systems, relatively little is known about this in humans so far.
Etiology
More than 60 genes have been identified as required for normal left-right axis specification, left-right patterning, or respiratory ciliary function. [19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43]
See the image below.
 Genes required for proper left-right asymmetry are shown. Genes are presented in five columns, according to the developmental phase in which they are currently thought to function. The leftmost column has the earliest functioning genes. The second column has genes required for the development of the node (or its equivalent). The third and fourth column have genes that are required for normal node cilia function. Genes in white, green, or blue denote those in which the proof came from studies of fruit fly (Drosophila melanogaster), zebrafish (Danio rerio), or frog (Xenopus laevis), respectively. Genes in brown are those studied in mice (Mus musculus), whereas those discovered in humans (Homo sapiens) are shown in red.
Genes required for proper left-right asymmetry are shown. Genes are presented in five columns, according to the developmental phase in which they are currently thought to function. The leftmost column has the earliest functioning genes. The second column has genes required for the development of the node (or its equivalent). The third and fourth column have genes that are required for normal node cilia function. Genes in white, green, or blue denote those in which the proof came from studies of fruit fly (Drosophila melanogaster), zebrafish (Danio rerio), or frog (Xenopus laevis), respectively. Genes in brown are those studied in mice (Mus musculus), whereas those discovered in humans (Homo sapiens) are shown in red.
Epidemiology
United States data
The worldwide incidence of heterotaxy syndrome is reportedly 1 case per 10,000 births. [44] This closely approximates the findings of the Baltimore-Washington Infant Study, in which the incidence of cardiac malformations associated with abnormal laterality was estimated at 1.44 cases per 10,000 live births. [45]
The true prevalence of heterotaxy syndrome is unknown, because many patients, especially those with left atrial appendage isomerism or polysplenia, have sufficiently mild heart disease such that the underlying diagnosis of heterotaxy may not even be considered by the clinician.
Race-, sex-, and age-related demographics
Although no predilection based on race has been identified, two studies noted a male-to-female ratio of 2:1.
The age at presentation is largely dependen on the severity of the heart disease.
Prognosis
The vast majority of patients with heterotaxy syndrome who have cardiovascular phenotypes significant enough to warrant cardiac surgical palliation undergo staged reconstruction to create Fontan-type circulatory arrangements; all of these patients can be expected to need cardiac transplantation in the second or third decades of life. Whether patients with heterotaxy syndrome who have cardiovascular defects that can be managed without the Fontan procedure fare worse than comparable patients without heterotaxy is currently unknown.
In a retrospective, single-center study (1997-2014) of 35 children with heterotaxy syndrome, the reported survival was 83% over a median follow-up of 65 months. [46] Of the 12 patients with poor outcomes (34.3%), 6 died, 1 underwent cardiac transplantation, and 5 had a New York Heart Association heart failure classification above III.
-
The structure and function of cilia is shown here. (A) Most motile cilia are organized with 9 microtubule doublets surrounding a core pair of doublets (9+2 configuration). Outer dynein arms (green) and inner dynein arms (blue) are shown. Cilia on the cells of the ventral node in the normal mouse embryo have no core doublet (a 9+0 configuration) and were initially thought to be nonmotile; however, upon closer scrutiny, node cilia were seen to have a rotatory motion (600 rpm). [Figure A is from Hirokawa N, Tanaka Y, Okada Y. Left-right determination: involvement of molecular motor KIF3, cilia, and nodal flow. Cold Spring Harb Perspect Biol. Jul 2009;1(1):a000802 and is reprinted with permission of Cold Spring Harbor Press.] (B) lrd (left-right dynein), the protein (green) mutated by the iv mutation, is also known as DNAH11, DNAHC11, and DLP11. [Figure B is from the United States Department of Energy Genomes to Life Program.] (C) The rotatory cone of each cilium is tilted posteriorly. Hence, the cilia make a leftward swing at the fluid surface and a rightward swing at the cellular surface. Because more viscous drag is present at the cellular surface, the rightward sweep is less effective at generating fluid movement than is the leftward sweep. [Figure C is from Hirokawa N, Tanaka Y, Okada Y, Takeda S. Nodal flow and the generation of left-right asymmetry. Cell 2006; 125:33-45 and is reproduced with permission from Cell Press.] A = anterior; L = left; P = posterior; r = Right.
-
Three phases of elaboration of left-right (LR) asymmetry are shown. The first step consists of differentiating the left and right sides on the cellular level. This probably takes place by means of a chiral molecule. (A) A subset of the cells (yellow) of the fairly early embryo undergo this process. (B) Localized cellular asymmetry is propagated between cells to cause LR determinants to accumulate on one side of the embryonic midline, possibly by a process involving transport through gap junctions. These determinants would then induce cascades of factors in multicellular fields of the embryo. (C) Finally, the asymmetric presence of these factors induces or suppresses asymmetrically located organs such as the spleen and regulates asymmetric morphogenesis of other organs such as the heart tube. Courtesy of Levin M, Mercola M. The compulsion of chirality: toward an understanding of left-right asymmetry. Genes Dev. Mar 15 1998;12(6):763-9.
-
Genes required for proper left-right asymmetry are shown. Genes are presented in five columns, according to the developmental phase in which they are currently thought to function. The leftmost column has the earliest functioning genes. The second column has genes required for the development of the node (or its equivalent). The third and fourth column have genes that are required for normal node cilia function. Genes in white, green, or blue denote those in which the proof came from studies of fruit fly (Drosophila melanogaster), zebrafish (Danio rerio), or frog (Xenopus laevis), respectively. Genes in brown are those studied in mice (Mus musculus), whereas those discovered in humans (Homo sapiens) are shown in red.
-
Axial magnetic resonance image (MRI) of a case of heterotaxy with polysplenia. (A) The abdominal aorta (abd ao) is on the left side of the spine (S), as is the left-sided azygos (L Azy). Two right-sided spleens (spl) are visible. LHV = left hepatic vein; RHV = right hepatic vein. (B) A common atrioventricular valve (black unlabelled arrows) is markedly malaligned to the right ventricle (RV). A diminutive left atrium (LA) is represented by only an appendage. The patient had an extracardiac conduit (EC) type of Fontan operation. No fenestration is noted between the EC and the neo-left atrium (neoLA). (C) Because this patient had subaortic stenosis, a proximal pulmonary artery-to-ascending aortic anastomosis was performed early in life, along with augmentation of the aortic arch. The L Azy connects to the left superior vena cava (LSVC). LU DAo = left upper descending aorta; Prox = proximal. (D) The LSVC connected originally to the coronary sinus (CS) and then to the right atrium. Despite the fact that the LSVC has been disconnected from the heart and anastomosed end-to-side to the left pulmonary artery, the CS remains large. The narrowed left ventricular outflow tract (LVOT) is seen. Ao = aorta; PA = pulmonary root; RLL PV = right lower lobe pulmonary vein. (E) Because this patient had absence of the hepatic segment of the inferior vena cava, the left-sided SVC-to-left pulmonary artery (LPA) anastomosis is referred to a left-sided Kawashima (LK). The anastomosis of the right superior vena cava to the right pulmonary artery is a right-sided bidirectional Glenn (R BDG) shunt. (F) The left lower lobe pulmonary vein (LLL PV), as part of this patient's totally anomalous pulmonary venous connection, connects to the original right atrium, which is now the neoLA.
-
Coronal magnetic resonance image (MRI) of the same patient as shown in the previous image. (A) Both superior vena cava (SVC)–to–pulmonary artery (PA) anastomoses can be seen. LCCA = left common carotid artery. (B) Three-dimensional surface rendering. RIA = right innominate artery. (C) Three-dimensional reconstruction of only the systemic venous pathway.
-
Malrotation of the gut. This upper gastrointestinal (GI) barium study of the same heterotaxy patient as shown in the previous two images shows a right-sided stomach (St), opposite of the normal site. The duodenum heads to the left, the duodenal-jejunal junction is to the left of the spine (opposite to what would be expected for situs inversus totalis), and the jejunum (J) stays left-sided.





